근육은 체중의 약 40%를 차지하며, 신체운동, 영양소 저장, 관절 안정화 등의 중요한 역할을 한다. 근육 기능이 저하되면 고관절 골절과 같은 신체적 장애가 증가하며 당뇨병, 지방간질환, 심혈관질환 등의 질병 위험도 높아진다(Kim and Kim, 2021). 현재 테스토스테론 보충제나 선택적 안드로겐 수용체 조절제 등으로 근감소증을 개선하려는 시도가 있지만(Dennison et al., 2017), 근육의 질을 결정하는 다양한 요인, 예를 들어 근섬유 유형, 근육 구조, 유산소 능력, 지방 축적 등에 대한 심층적인 연구는 부족한 실정이다(McGregor et al., 2014).
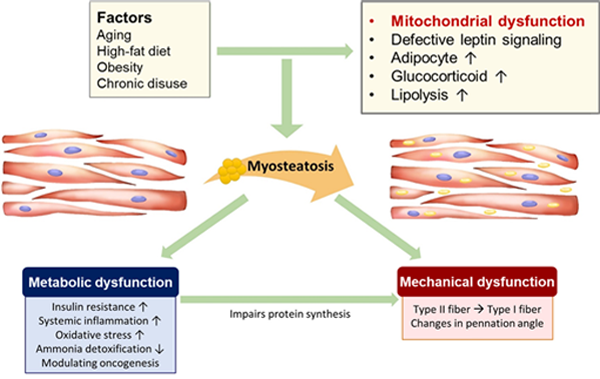
근지방증(myosteatosis)은 근육 내에 지방이 과도하게 축적되어 근육의 질이 저하된 상태를 말한다. 근육과 지방조직에서 지질 대사 능력이 과포화되었을 때 근육 내에 지방이 축적되고, 또한 나이가 들수록 근육 내 지방량이 증가하는 것으로 알려져 있다. 고지방식이, 운동 부족, 내분비 이상 역시 근지방증의 주요 유발 요인이다(Henin et al., 2024). 근지방증의 발생기전으로 렙틴 신호전달(leptin signaling) 결함, 근육줄기세포 또는 섬유아디포세포 전구세포(fibroadipogenic precursor cell)의 관여, 신경근의 변화, 미토콘드리아 기능 장애 등이 제시되고 있다(Correa-de-Araujo et al., 2020).
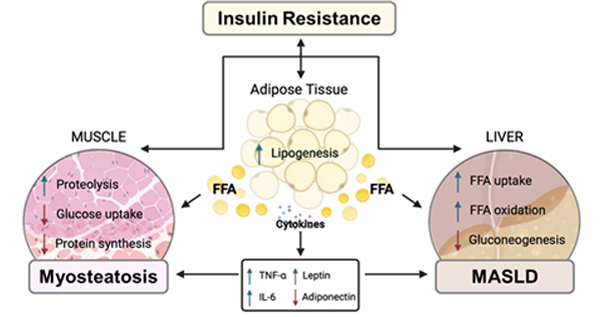
골격근 내에서 지질은 (1) 근간 지방조직(intermuscular adipose tissue, IMAT): 근막 아래와 근육 사이에 위치하는 세포 외 지방조직, (2) 근내 지방조직(intramuscular adipose tissue): 개별 근섬유 내에 위치하는 세포 외 지방조직, (3) 근세포 내 지질(intramyocellular lipids, IMCL)의 형태로 존재한다(Correa-de-Araujo et al., 2020). IMCL은 제2형 당뇨병 환자의 인슐린 저항성을 악화시키고, IMAT은 대사 증후군 및 노화에 따른 신체 기능 저하와 관련이 있다(Goodpaster et al., 2023; Laurens and Moro, 2016).
과거 근지방증은 근감소증의 부수적인 질환으로 여겨졌으나, 현재는 대사질환에 영향을 미치는 독립적 위험 요인으로 간주되어 관련 연구가 활발히 진행되고 있다. 근지방증은 노화와 대사질환에서 다양한 부정적 임상 결과와 연관이 있으며, 특히 대사이상지방간질환(MASLD) 발생 및 중증도에 영향을 줄 수 있는 독립적인 원인으로 밝혀지고 있다(Henin et al., 2024). 따라서, 근지방증 치료가 전신적 대사질환 개선에 효과적일 것으로 기대하고 있으나 예후적 영향 및 치료 전략에 관한 대규모 연구는 아직 부족하다.
그림 1. 근육, 간, 지방조직 간의 상호작용 (Marco et al., 2022)
근지방증을 유발하는 핵심 원인: 근육세포의 미토콘드리아 기능 상실
근육은 다양한 근섬유 유형으로 구성된 조직으로, 크게 type 1 ‘느린 수축’ 섬유와 type 2 ‘빠른 수축’ 섬유로 구분된다. 빠른 수축 섬유는 서로 다른 MyHC 유전자 발현에 따라 2a, 2x, 2b로 세분화된다. Type 1과 type 2a 섬유는 미토콘드리아 수가 많고 주로 산화 대사를 이용하는 반면, type 2x와 type 2b 섬유는 미토콘드리아 수가 제한적이며 주로 해당작용을 통해 에너지를 생산한다(Schiaffino and Reggiani, 2011).
미토콘드리아의 정상적 기능은 근육 내 지질의 산화 대사에 필수적이며 미토콘드리아의 기능 장애는 근육세포에서 산화 스트레스, 지질 독성, 염증과 밀접하게 연관되어 있다. 미토콘드리아 생합성은 여러 미토콘드리아 호흡 복합체 단백질과 미토콘드리아 전사 인자 A(TFAM)의 발현을 조절하는 NRF1, GA 결합 단백(GABP)에 의해 조절되며 NRF1과 GABP의 기능은 이들의 공동 활성인자인 PGC-1α와의 상호작용에 의해 증대된다(Dominy and Puigserver, 2013). 근지방증 마우스 모델의 근육에서 NRF1과 TFAM의 발현이 감소하였으며 비만 및 당뇨병 환자의 근육에서도 중성지방 축적과 함께 PGC-1α에 의해 조절되는 미토콘드리아 유전자 발현이 감소되었다(Gumucio et al., 2019; Roden, 2005). 이러한 관찰들은 근육의 미토콘드리아 기능이 근육 내 지방 감소 및 근육의 질적 개선에 중요한 역할을 한다는 것을 시사한다.
그림 2. 근지방증의 병태생리 (Ahn et al., 2021)
근육세포의 미토콘드리아 표적 인자, 핵 수용체 RORα
RORα는 스테로이드 호르몬 수용체 패밀리에 속하는 리간드 의존성 전사 인자로서 발달, 대사, 저산소증 반응 등과 관련된 유전자의 발현을 조절한다. 우리 연구실에서는 RORα가 간세포에서 미토콘드리아의 역동성과 미토콘드리아 선택적 오토파지를 조절하여 미토콘드리아의 기능을 증진하고 대사이상지방간질환을 개선한다는 것을 보고한 바 있다(Kim et al., 2012; Kim et al., 2017; Kim et al., 2021).
RORα의 근육 내 잠재적 역할은 RORα 결손 staggerer 마우스에서 근 위축이 나타남으로써 처음으로 제시되었으며, RORα의 활성이 근육 내 지질 항상성 관련 다수 유전자의 발현과 연관되어 있음이 보고된 바 있다. 따라서, 근육 내 RORα를 활성화하여 근육을 질적으로 향상할 수 있다는 가설을 수립하였으며, 대사이상 지방간 질환이 동반된 근지방증 마우스 모델을 사용하여 근육에서 RORα의 역할을 규명하고, RORα가 미토콘드리아 기능을 조절하는 분자기전을 이해하고자 하였다.
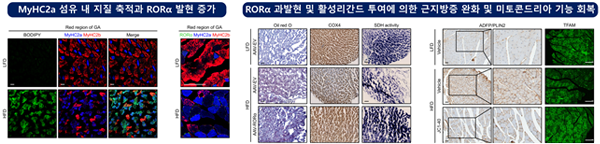
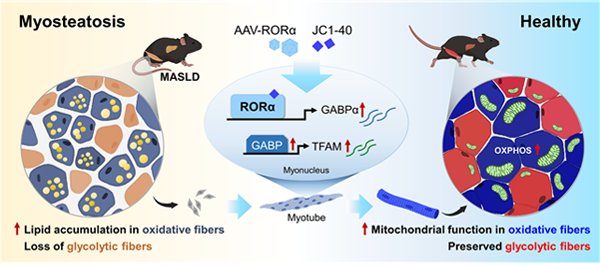
고지질식이로 유도된 근지방증 마우스의 gastrocnemius 적색 영역에서 산화성 MyHC2a 섬유의 수가 증가하는 것을 확인하였으며 RORα는 MyHC2a 섬유 주변에서 발현이 증가되어 있었다. RORα 리간드인 cholesterol sulfate와 JC1-40 처리에 의하여 MyHC1 및 MyHC2a 섬유의 수가 증가하였고, 지방산 존재 하에서 MyHC2a 섬유 내 지질 축적이 감소하였다. RORα 리간드 처리는 미토콘드리아 막전위와 산소소비량을 증가시켰으며 분자기전으로서 RORα가 GABP 유전자의 프로모터에 결합하여 TFAM 전사 발현을 증가시킨다는 것을 밝혔다. RORα 근육 내 과발현은 지질 함량을 감소시키고, TFAM 발현과 미토콘드리아 기능을 회복시켰다. 또한 JC1-40를 경구투여하였을 때 마우스 근육 내 지질 축적이 감소되었으며 미토콘드리아 단백질의 발현이 회복되었다. 이들 결과는 RORα가 산화성 섬유 내 미토콘드리아의 양과 산화 능력을 조절하여 근지방증을 개선함을 알려주며 본 연구를 통해 RORα 활성제를 이용한 미토콘드리아 표적 근지방증 치료 전략을 제시하였다.
그림 3. RORα-GABP-TFAM axis에 의한 근지방증 개선 효과 (Kim et al., 2024)
맺음말
근육은 다양한 근섬유의 조합으로 이루어져 있으며, 각 섬유는 고유한 수축 특성과 대사적 표현형을 가지고 있다. 이들 섬유는 미토콘드리아 양의 차이로 명확히 구분되지만, 양적 차이 외에도 근육 기능 최적화를 위해 중요한 지질 산화 능력, 활성산소종 관련 신호체계, 항산화 능력 등의 미토콘드리아 표현형의 차이도 나타난다. 또한, 다양한 병태생리학적 요인들에 복잡하게 반응하는 근육 내 여러 세포 집단에서 근지방증의 인과적 경로를 규명하는 일은 매우 도전적이다. 올해 발표한 근지방증 연구(Kim et al., Journal of Cachexia, Sarcopenia and Muscle, 2024)는 근육 내 병태생리 현상을 탐구하는 첫 시도이며 앞으로 근육 내 지방조직의 기원, 근지방증 발생과 관련한 세포 및 조직 간 상호작용, 섬유별 미토콘드리아 표현형에 대한 포괄적인 연구가 활발히 일어나기를 기대한다.
참고문헌
Ahn, H., Kim, D.W., Ko, Y., Ha, J., Shin, Y.B., Lee, J., Sung, Y.S., and Kim, K.W. (2021). Updated systematic review and meta-analysis on diagnostic issues and the prognostic impact of myosteatosis: A new paradigm beyond sarcopenia. Ageing Res Rev 70, 101398.
Correa-de-Araujo, R., Addison, O., Miljkovic, I., Goodpaster, B.H., Bergman, B.C., Clark, R.V., Elena, J.W., Esser, K.A., Ferrucci, L., Harris-Love, M.O., et al. (2020). Myosteatosis in the Context of Skeletal Muscle Function Deficit: An Interdisciplinary Workshop at the National Institute on Aging. Front Physiol 11, 963.
Dennison, E.M., Sayer, A.A., and Cooper, C. (2017). Epidemiology of sarcopenia and insight into possible therapeutic targets. Nat Rev Rheumatol 13, 340-347.
Dominy, J.E., and Puigserver, P. (2013). Mitochondrial biogenesis through activation of nuclear signaling proteins. Cold Spring Harb Perspect Biol 5.
Gumucio, J.P., Qasawa, A.H., Ferrara, P.J., Malik, A.N., Funai, K., McDonagh, B., and Mendias, C.L. (2019). Reduced mitochondrial lipid oxidation leads to fat accumulation in myosteatosis. Faseb j 33, 7863-7881.
Henin, G., Loumaye, A., Leclercq, I.A., and Lanthier, N. (2024). Myosteatosis: Diagnosis, pathophysiology and consequences in metabolic dysfunction-associated steatotic liver disease. JHEP Rep 6, 100963.
Kim, E.-J., Yoon, Y.-S., Hong, S., Son, H.-Y., Na, T.-Y., Lee, M.-H., Kang, H.-J., Park, J., Cho, W.-J., Kim, S.-G., et al. (2012). Retinoic acid receptor–related orphan receptor α–induced activation of adenosine monophosphate–activated protein kinase results in attenuation of hepatic steatosis. Hepatology 55.
Kim, H.-J., Han, Y.-H., Na, H., Kim, J.-Y., Kim, T., Kim, H.-J., Shin, C., Lee, J.W., and Lee, M.-O. (2017). Liver-specific deletion of RORα aggravates diet-induced nonalcoholic steatohepatitis by inducing mitochondrial dysfunction. Scientific Reports 7, 16041.
Kim, H.J., Han, Y.H., Kim, J.Y., and Lee, M.O. (2021). RORα Enhances Lysosomal Acidification and Autophagic Flux in the Hepatocytes. Hepatol Commun 5, 2121-2138.
Kim, H.K., and Kim, C.H. (2021). Quality Matters as Much as Quantity of Skeletal Muscle: Clinical Implications of Myosteatosis in Cardiometabolic Health. Endocrinol Metab (Seoul) 36, 1161-1174.
Kim, H.J., Lee, S.H., Jeong, C., Han, Y.H., and Lee, M.O. (2024). RORα-GABP-TFAM axis alleviates myosteatosis with fatty atrophy through reinforcement of mitochondrial capacity. J Cachexia Sarcopenia Muscle 15, 615-630.
Laurens, C., and Moro, C. (2016). Intramyocellular fat storage in metabolic diseases. Hormone Molecular Biology and Clinical Investigation 26, 43-52.
Marco, A., Claudio, C.-V., Juan Pablo, A., Francisco, B., Rene, B., Francisco, J.G., Iram, G., and Daniel, C. (2022). Sarcopenia in the setting of nonalcoholic fatty liver. Metabolism and Target Organ Damage 2, 2.
McGregor, R.A., Cameron-Smith, D., and Poppitt, S.D. (2014). It is not just muscle mass: a review of muscle quality, composition and metabolism during ageing as determinants of muscle function and mobility in later life. Longev Healthspan 3, 9.
Roden, M. (2005). Muscle triglycerides and mitochondrial function: possible mechanisms for the development of type 2 diabetes. Int J Obes (Lond) 29 Suppl 2, S111-115.
Schiaffino, S., and Reggiani, C. (2011). Fiber types in mammalian skeletal muscles. Physiol Rev 91, 1447-1531.

 대한내분비학회 지난 호 웹진 보기
대한내분비학회 지난 호 웹진 보기